

Research
Section Mammalogy
My research interests centre around vertebrate functional morphology, and evolutionary adaptations. I primarily use palaeophenomics in the form of geometric morphometrics to assess evolutionary adaptations through time and space, as well as bone microstructural analyses and finite element analyses. A few important examples are highlighted below.
Evolutionary Adaptations throughout the Quaternary
One of my long term scientific goals is to understand insular evolution and the processes involved. This is best achieved using a cross-disciplinary approach to analyse size, morphology and development of island animals on all organismal levels. My projects so far have focused on various vertebrates using a range of analytical techniques to increase understanding of the combined influence of isolation in a mammalian predator-free environment.
In the light of current environmental changes, Pleistocene animals are an exceptionally good study system. Ice age ecosystems went through global warming at the Pleistocene-Holocene boundary and, whereas some animals managed to adapt, others went (locally) extinct. Furthermore, Pleistocene animals are very closely related to extant animals, sometimes even belonging to the same species. Understanding how and why these animals adapted is paramount for understanding and helping extant animals.
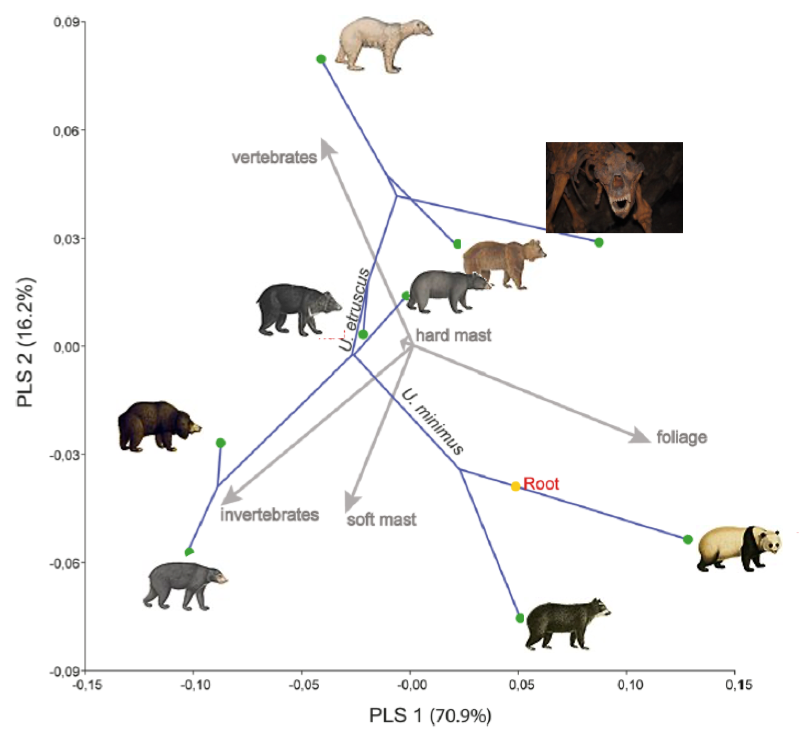
Amongst others, I use bears as a model species. In the Pleistocene, there were two bear taxa in Europe: various cave bear lineages (Ursus spelaeus s.l.) and the brown bear (Ursus arctos). It is imperative that we understand what caused the cave bears to perish in the context of climatic warming at the end of the Pleistocene, whereas the brown bear survived, to prevent extant bear species from meeting the same fate. Until recently, the diet and evolution of the cave bear were largely unknown. I have analysed cave bears and their living relatives using palaeophenomics. The cave bears shows a similar evolutionary trajectory through morphospace to the panda (Ailuropoda melanoleuca) indicating similar adaptations.
Evolution on Islands – Nature’s Laboratory
It may be expected that some animals are more and others less prone to morphological adaptations brought on by environmental changes. For example, insular animals during the Cenozoic have been shown to have undergone many, and sometimes profound, morphological changes in their new environments. Understanding evolution on islands is imperative for understanding potential changes in animal adaptations in relation to current climatic changes. Nowhere else has nature provided us with the opportunity to test evolutionary hypotheses pertaining to adaptations and functional morphology in duplo or triplo (which would be the norm in a chemistry lab), making islands the only habitats in the world where common and taxon-specific evolutionary adaptations can be distinguished, quantified, and statistically verified using palaeophenomics.
An interesting example of insular dwarfing is Homo floresiensis. In my research, I compare H. florsiensis with mammalian insular dwarfs. H. floresiensis displays adaptations to low gear locomotion. Furthermore, H. floresiensis displays archaic wrist morphology and heterochronic features in the skull and postcranial remains. These are features common to many endemic island taxa.
Another endemic hominin is Homo luzonensis from the Philippines. Not much is known about this species, but they were small in stature, like H. floresiensis, and displayed a mixture of archaic and derived morphologies. Our research is the first to use synchrotron radiation X-ray tomography to assess developmental speed in fossils. Our research has shown that H. luzonensis probably had a developmental pattern that was quite similar to modern apes.

The dodo (Raphus cucullatus) might be the most enigmatic bird of all times. I am a member of the Dodo Research Programme and I have contributed to new knowledge regarding the dodo and its environment. Amongst others, I have provided an estimate of the dodo’s mass based on the largest sample of dodo femora ever measured. The results indicate that the mean mass of the dodo was circa 12 kg, which is approximately five times as heavy as the largest living pigeons and doves, to which the dodo belongs.
Haast’s eagle (Hieraaetus (=Harpagornis) moorei) was the largest eagle to have ever existed and once lived in New Zealand. DNA analysis has demonstrated that its closest living relative is one of the world’s smallest eagles, the little eagle (Hieraaetus morphnoides). Haast’s eagle was more than an order of magnitude heavier than the the little eagle and represents an extraordinary example of rapid evolution within less than 2 million years. Based on geometric morphometrics and finite element analysis, we concluded that Haast’s eagle is characterized by a unique mosaic of eagle and vulture-like features that probably evolved in response to highly demanding selective pressures to routinely hunt and eat particularly large prey in order to maintain their giant size.
Phenomics
Preliminary microstructure analysis of European squirrel (Sciurus vulgaris fuscoater) humeri, all from Bavaria (Germany), that died in different seasons in a short 10-year period between 1907 and 1917, were analysed using a largely automated protocol for data acquisition. Analyses showed highly significant differences in bone volume between the seasons, which were strongly suggestive of intraindividual changes through time, possibly caused by a lack of dietary calcium in winter. This was previously unknown to science. This project serves as a proof of concept for palaeophenomics, as a potent and versatile framework to robustly represent morphology, showing the potential of big data in virtual palaeobiology.
